Paleodieta y movilidad: análisis isotópicos en restos óseos humanos de la cueva de La Sepultura (3050-2850 a.P.) sierra de Naola Tula, Tamaulipas
Jesús Ernesto Velasco González
Tonantzin Silva Cárdenas
Carlos Vanueth Pérez Silva
Instituto Nacional de Antropología e Historia (INAH), Tamaulipas, México,
correos electrónicos: afernestovelasco@gmail.com, tonantzinsilva@hotmail.com, pedernal7501@gmail.com
Abigail Meza Peñaloza
Carlos Serrano Sánchez
Instituto de Investigaciones Antropológicas, Universidad Nacional Autónoma
de México (UNAM), Ciudad de México, México,
correos electrónicos: abigailm@unam.mx, cserrano@unam.mx
Francisco Javier Otero Trujano
Edith Cienfuegos
Instituto de Geología, Universidad Nacional Autónoma de México (UNAM),
Ciudad de México, México,
correos electrónicos: fjavierotero@yahoo.com.mx, edithca@unam.mx
José Antonio Caro Gómez
Universidad de Córdoba, España,
correo electrónico: gt1cagoj@uco.es
Genaro Álvarez García
Sociedad Espeleológica GEOS, España,
correo electrónico: segeos@telefonica.net
Recibido el 12 de enero de 2022; aceptado el 10 de febrero de 2022
Resumen: La aplicación de técnicas isotópicas en huesos y dientes humanos orientadas a la reconstrucción de las dietas del pasado (Ambrose y De Niro, 1986; De Niro, 1985; Kellner y Schoeninger, 2007; Lee-Thorp et al., 2008; Morales Puente et al., 2012; Schwarcz, 2000; Shoeninger y De Niro, 1983; Van der Merwe et al., 1978; Vogel et al., 1977), han demostrado su importancia en ampliar el registro arqueológico en temas como la domesticación, dispersión y uso del maíz (Eubanks, 2001; Flannery, 1973; Harlan, 1971; Hole, 1993; Mangelsdorf et al., 1964; William et al., 2009) como de otras plantas cultivables y recolectadas. Adicionalmente, también se han ido incorporando otros aspectos de interés antropológico, entre los que se encuentra el origen y movilidad geográfica de las personas (Daux et al. 2008; Delgado et al., 1995; Gat, 1996). Desde la lógica bioquímica del fraccionamiento isotópico en colágeno y bio-apatita (Jim et al., 2004; Krueger y Sullivan, 1984; Lee-Thorp et al., 1989), el modelo simple de regresión lineal o bivariado (Froehle et al., 2010; Kellner y Schoeninger, 2007), junto con el de funciones discriminantes o multivariado (Froehle et al., 2012), brindan la posibilidad de contrastar dietas con firmas isotópicas similares, proporcionando una base confiable de diferenciación individual y poblacional sobre las fuentes de proteína específicas sean éstas de origen vegetal y animal terrestre, lacustre o marina. Lo anterior es útil para la explicación de los procesos de obtención, producción y consumo de alimentos, ofreciendo datos cuantitativos para el entendimiento de las dietas mixtas en entornos concretos y contextos de interface entre poblaciones de cazadores-recolectores-pescadores, como en la de agricultores de baja y alta intensidad (Hard y Katzenberg, 2011; Smith, 1998a, 2001).
El análisis isotópico de carbono (13C) en colágeno y bioapatita de 20 muestras obtenidas de hueso y dientes humanos procedentes de la cueva de la Sepultura 3050-2850 a.P. (1400-1100 a.C.) (Caro et al., 2012; Pérez Silva et al., 2012a; Pérez Silva et al., 2012b; Pérez Silva et al., 2011a; Pérez Silva et al., 2011b; Silva et al., 2013a; Silva et al., 2013b; Silva González et al., 2013c; Valdiosera, 2016; Velasco González, 2011, 2016; Velasco González et al., 2013), confirma la aseveración de que durante la fase Mesa del Guaje 3600-3000 a.P. (1650-1050 a.C), de la secuencia cultural establecida para el suroeste de Tamaulipas (Hanselka, 2008, 2011; MacNeish, 1958, 1998, 2001), se tiene el consumo de plantas C3 con una importante dominancia de plantas CAM y C4. Encontrando además en el presente estudio que, los altos niveles de nitrógeno (15N) en todas las muestras analizadas, indican una dieta integrada por animales terrestres y probablemente lacustres, ribereños o marinos. Lo anterior plantea el uso de una amplia y rica variedad dietética producto no solo de cierta movilidad restringida, sino de posibles desplazamientos efímeros en un medio ambiente de transición ecológica entre la sierra y la costa, según lo insinúa, el análisis de isótopos de oxígeno (18O). Dado lo anterior, se proporcionan nuevos datos al problema histórico-cultural y conceptual, sobre el incremento del uso del maíz y otras plantas cultivables durante el desarrollo e interacción de los grupos cazadores-recolectores-pescadores, y agricultores de baja intensidad entre el Altiplano del norte, la Sierra Madre Oriental y la Planicie costera del golfo de México durante el Formativo mesoamericano.
Palabras clave: paleodieta, movilidad, isótopos estables, colágeno, bioapatita, caza-recolección-pesca-agricultura.
Paleodiet and mobility: isotopic analysis in human bone remains from the Sepultura cave (3050-2850 b.C.) sierra de Naola Tula, Tamaulipas
Abstract: The isotope techniques applied in human bones and teeth aimed to reconstruct the diets of the past (Ambrose & De Niro, 1986; De Niro, 1985; Kellner & Schoeninger, 2007; Lee-Thorp et al., 2008; Morales Puente et al., 2012; Schwarcz, 2000; Shoeninger & De Niro, 1983b; Van der Merwe et al., 1978; Vogel et al., 1977), demonstrate the importance to extend topics on domestication, dispersal and use of maize (Eubanks, 2001; Flannery, 1973; Harlan, 1971; Hole, 1993; Mangelsdorf et al., 1964; William et al., 2009), as others cultivated and recollected plants in the archaeology context. In addition, they incorporate other aspects from anthropology interests such the origin and geographical mobility of the people (Daux et al., 2008; Delgado et al., 1995; Gat, 1996). From in the biochemical logic of isotopic fractionation in collagen and bio-apatite (Jim et al., 2004; Krueger & Sullivan, 1984; Lee-Thorp et al., 1989), the linear simple or bivariate regression model (Froehle et al., 2010; Kellner & Schoeninger, 2007), together discriminant function analysis or multivariate model (Froehle et al., 2012), provide the possibility to contrasts diets with similar isotopic signatures, offering a confiability basis to individual and population differences about sources of specific protein whether they are vegetal or animal terrestrial, lacustrine or marine resources. The above is helpful from explain the processes to produce, obtain and consume foods, offering cuantitaive dataset to the understand the mixed diets in ecological and interface contexts between hunter gatherers or low level farmers and intensive agriculturalists (Hard & Katzenberg, 2011; Smith, 1998a, 2001).
The isotopic analysis for carbon (13C) in collagen and apatite from 20 samples obtained from bone and teeth in the Sepulture cave 3050-2850 B.P. (Caro et al., 2012; Pérez Silva et al., 2012a; Pérez Silva et al., 2012b; Pérez Silva et al., 2011a; Pérez Silva et al., 2011b; Silva González et al., 2013a; Silva et al., 2013b; Silva et al., 2013c; Valdiosera, 2016; Velasco González, 2011, 2016; Velasco González et al., 2013), confirms that in the Mesa del Guaje phase 3600-3000 B.P. from the cultural sequence established for the Southwest of Tamaulipas (Hanselka, 2008, 2011; MacNeish, 1958, 1998, 2001), they´re had the consume for C3 plants with and important prevalence for CAM/C4 plants. In addition, in this study we found which the higher level for nitrogen (15N) in all analyzed samples, indicate a diet integrated for terrestrial and probably lacustrine and marine animals. This arise from the use for a wide and rich variety of diet result no only some restrained mobility, otherwise the ephemeral possibly displacements in an ecology transition from mountains and the coast, as hinted the oxygen isotope (18O) analysis. Given the above, we proportionate new data to the cultural-historic and conceptual problem, about the increase use of maize and the other cultivate plants during the interactive and evolution of hunter-gatherer and low level farmers groups between north plateau, Sierra Madre Oriental and the coastal gulf plain from Mexico in the Formative Mesoamerican period.
Key words: paleodiet, mobility, stable isotopes, collagen, bio-apatite, hunter-gatherer-fisher-agriculturalist.
Introducción
A diferencia de las grandes llanuras del sur de Texas y noreste de México en las que no existen cordilleras, la planicie costera del golfo en Tamaulipas, se encuentra interrumpida por dos elevaciones importantes: la sierra de Tamaulipas y la sierra Madre Oriental (Salinas Rivera, 2012). Esta variedad fisiográfica integra una gran riqueza en biodiversidad, aunque también, una profusa herencia histórico-cultural con la presencia de numerosos sitios arqueológicos e históricos que hacen referencia tanto a complejos de cazadores-recolectores-pescadores, como también de poblaciones semi agrícolas y agrícolas de la época prehistórica y época del contacto europeo (Ekholm, 1943, 1944; Hughes, 1947; Krieger, 1945; MacNeish, 1947, 1949, 1954; Stresser-Pean, 1977). Es decir, dicha área se caracteriza no solamente por la problemática etnohistórica de la interacción de sociedades de cazadores recolectores y agricultores, sino también por la de los horticultores parciales o de baja intensidad (Kirchhoff, 1943a, 1943b, 1954; Smith, 1997, 1998a, 1998b, 2001), involucrando un escenario particular y un nexo común para el estudio de las economías mixtas entre la denominada “frontera” septentrional de Mesoamérica y el Norte de México (Figura 1).
En este contexto, investigaciones arqueológicas y de antropología física en la cueva de Las Calaveras o La Sepultura, localizada en la sierra de Naola, estribación de la sierra Madre Oriental en el municipio de Tula al suroeste de Tamaulipas (Caro Gómez et al., 2012; Pérez Silva et al., 2011a; Pérez Silva et al., 2011b; Velasco González, 2016; Velasco González et al. 2013) es sugerente, pues su estudio, arroja información sólida que ayuda a examinar el problema referido en párrafos anteriores. Dicho sitio, se emplaza sobre un abanico aluvial que forma parte de un sistema de cuencas endorreicas con vegetación propia

Figura 1. Ubicación del área de estudio. Tomado de Kirchhof (1954)
del desierto chihuahuense a 1630 msnm (Pérez Silva et al., 2011b). La entrada a la cueva es horizontal, tiene una abertura aproximada de metro y medio por un metro de altura, sin embargo, la cámara es de mayores dimensiones, ya que conforme se avanza hacia el interior el suelo va en descenso
(Figura 2) aumentando la altura de la bóveda que presenta forma de catedral (Caro Gómez et al., 2012). Aunque la superficie es en mayor parte rocosa, algunas secciones muestran acumulación de sedimento por acción erosiva, como lo es cerca del acceso y a unos 7 m de profundidad. En este espacio (Figura 3) se localizaron en superficie cuatro conjuntos principales de restos
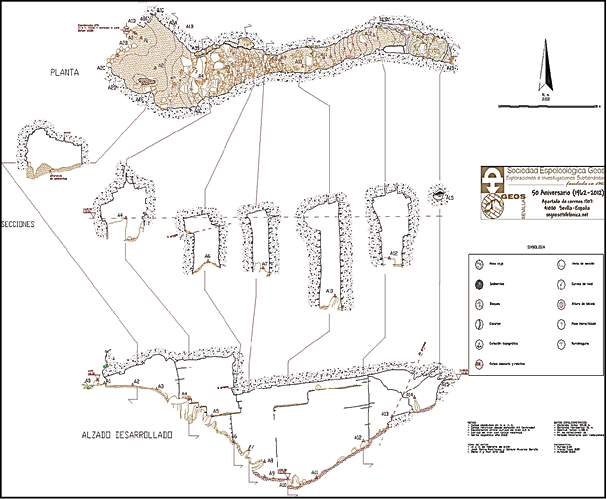
Figura 2. Plano de la cueva de La Sepultura. Tomado de Caro Gómez et al. (2012)
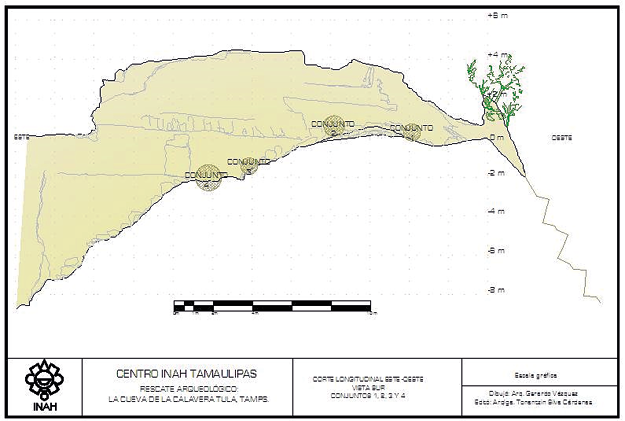
Figura 3. Ubicaciones principales de restos óseos humanos en superficie
(Pérez Silva et al., 2010)
óseos humanos desarticulados y mezclados entre sí (Pérez et al., 2010). No obstante, durante el proceso de excavación arqueológica, se identificaron otra gran cantidad de huesos dislocados, en adición a objetos de lítica tallada, concha, hueso, cestería, cordelería, petates y semillas de algunas plantas, incluyendo granos de maíz (Adriano-Morán y Martínez Yrizar et al., s.f.; Pérez Silva et al., 2011a). No se halló evidencia alguna de cerámica ni figurillas de barro, pero dieciséis fechas por radiocarbono AMS (Tabla 1) y el perfil osteológico correspondiente, apuntan a que el sitio tuvo una ocupación funeraria afín a la tradición del Desierto entre 3050-2850 a.P. (1400-1100 a.C.) (Velasco González, 2011, 2016).
Tabla 1. Fechamientos absolutos 14C (AMS) de la cueva de La Sepultura o Calaveras
|
Lab. Code |
Sample |
Site |
Date δ13C BP |
2 σ 95% CalDate |
|
Beta - 457995 |
md12 |
TCS-1 |
2850+30 |
1110 - 925 BC |
|
LUR:UNAM-1216 |
or-2 |
TCS-1 |
2910±70 |
1313 - 914 BC |
|
Beta - 473644 |
md19 |
TCS-1 |
2940+30 |
1256 - 1043 BC |
|
Beta - 473645 |
md20 |
TCS-1 |
2950+30 |
1259 - 1050 BC |
|
Beta - 473639 |
md13 |
TCS-1 |
2970+30 |
1073 - 1057 BC |
|
Beta - 473640 |
md14 |
TCS-1 |
2980+30 |
1371 - 1111 BC |
|
Beta - 473641 |
md15 |
TCS-1 |
2980+30 |
1371 - 1111 BC |
|
Beta - 473642 |
md16 |
TCS-1 |
2980+30 |
1371 - 1111 BC |
|
Beta - 473643 |
md17 |
TCS-1 |
2990+30 |
1373 - 1117 BC |
|
Beta - 401542 |
A1-2 |
TCS-1 |
3000±30 |
1370 - 1125 BC |
|
Beta - 457996 |
md21 |
TCS-1 |
3000+30 |
1370 - 1125 BC |
|
Beta - 457997 |
md22 |
TCS-1 |
3000+30 |
1370 - 1125 BC |
|
Beta - 457998 |
md23 |
TCS-1 |
3010+30 |
1380 - 1160 BC |
|
Beta - 401543 |
B2-1 |
TCS-1 |
3020±30 |
1385 - 1130 BC |
|
OxA-26054 |
N1 |
TCS-1 |
3020±26 |
1387 - 1134 BC |
|
Beta - 401541 |
A1-1 |
TCS-1 |
3050±30 |
1405 - 1220 BC |
Planteamiento
La secuencia cultural del suroeste de Tamaulipas (MacNeish, 1958, 1998, 2001), establece que durante la fase Mesa del Guaje (1650-1050 a.C.), se manifiesta la intensificación de plantas cultivables como el maíz en combinación con la recolección y la caza, así como un incremento de la vida sedentaria paralela a áreas adyacentes desarrolladas hacia la costa del golfo desde el 1600 a.C. (Hanselka, 2008, 2011; MacNeish, 1947, 1950, 1958; Merino Carrión y García Cook, 1987, 1997, 2002, 2004). Para la Zona Media Potosina, esta se ve descrita hacia el final de la fase Venadito I (1000-200 a.C.), periodo en el que sociedades de agricultores se desarrollan en un área de contacto asociada a cazadores-recolectores (Rodriguez Loubet, 1985; Tesch, 1991, 1996). Estos procesos pueden abordarse a través del análisis de isótopos estables de carbono en colágeno (δ13Ccol) y bio-apatita (δ13Cap) de huesos y dientes humanos, identificando a su vez, las proporciones en la dieta de la fuente de proteína individual y poblacional no solo a nivel de desplazamiento isotópico (13C/15N), sino de manera específica, a partir de considerar los distintos fraccionamientos que derivan de otros procesos fisiológicos como los del metabolismo (Jim et al., 2004; Krueger y Sullivan, 1984; Lee-Thorp et al., 1989; Lyne et al., 2001; Minagawa y Wada, 1984; Nier y Gulbransen, 1939; Shoeninger y De Niro, 1983, 1984; Shoeninger et al., 1983; Tieszen y Fagre, 1993).
Reconocer las fuentes de proteína total y directa ya sea de origen vegetal o animal, terrestre o acuático, es posible gracias a las diferentes rutas que siguen el carbono y nitrógeno en los tejidos, siendo de gran ayuda, para aquellos contextos donde la escasez de materiales arqueológicos es recurrente. De manera adicional, los isótopos estables de oxígeno (δ18O) en bio-apatita de los carbonatos, en relación con el agua meteórica δ2H de la región, son de gran ayuda para inferir patrones de movilidad geográfica, gracias a que este isótopo, se fija en el proceso de formación de los dientes durante el crecimiento y también varía en los procesos de recambio o remodelación del hueso (Daux et al., 2008; Delgado et al., 1995). Con lo anterior, podemos establecer aproximaciones detalladas sobre los procesos de adaptación alimentaria y movilidad, en un contexto de intensificación de plantas cultivables entre la zona del altiplano del norte, sierra Madre Oriental y la costa del golfo de México durante el Formativo.
Métodos, materiales y técnicas
Trabajos experimentales muestran que diferenciar los componentes energéticos por un lado y proteínas por otro solamente empleando datos absolutos (la diferencia aritmética entre los valores δ13Cap y δ13Ccol se denomina Δ 13Cap-col)
resulta poco fiable (Kellner y Schoeninger, 2007). El modelo simple de regresión lineal o bivariado, esquematiza mejor las fuentes de energía y proteína, ya que se ha descubierto que la mayoría de los átomos de carbono en el colágeno (δ13Ccol) proceden de las proteínas de la dieta (δ13Cdieta proteína), mientras el carbono contenido en los carbonatos o bio-apatita del hueso y esmalte (δ13Cap) proviene de lo que se ha denominado dieta total (δ13Cdieta total). Es decir, los hidratos de carbono, lípidos y proteína no utilizada en la síntesis de proteínas, puesto que la formación de la apatita del hueso se encuentra en equilibrio con el carbonato de la sangre que es en sí mismo un producto del metabolismo (Ambrose, 1993; Krueger y Sullivan, 1984; Shoeninger y De Niro, 1983; Tieszen y Fagre, 1993). Aplicado a la reconstrucción de la dieta en poblaciones humanas antiguas, la posición relativa de un individuo sobre la línea a lo largo del eje δ13Ccol muestra información isotópica sobre las fuentes directas de proteínas, mientras que la posición a lo largo del eje δ13Cap indicará la relación del conjunto de alimentos C3 a C4 y/o marina. Ni el tamaño del cuerpo ni la posición trófica parecen afectar estas relaciones (Froehle et al., 2010; Kellner y Schoninger, 2007). De este modo, evitando parte de la redundancia inherente a la separación apatita colágeno (Δ13Ccol-ap) derivado de la interpretación subjetiva de datos absolutos, se pueden obtener estimaciones más precisas a través de referencias basadas en modelos predictivos, pues dichas relaciones, se encuentran debidamente representadas a través de tres líneas de regresión que muestran constante las fuentes de proteína C3 o C4, mientras de forma variada el tipo de energía C3 a C4 y/o marina. Es decir, el extremo inferior de cada línea indica una dieta que consta de 100% de energía C3, mientras los extremos superiores representan una dieta total 100% de tipo C4 o marino (Kellner y Schoninger, 2007).
Aplicaciones de este modelo son las expuestas por Schwarcz (2000), quien afirma que en poblaciones humanas que viven con niveles bajos de ingesta de proteína, la composición isotópica del colágeno óseo tiende a comportarse conforme al modelo lineal mixto. En este tipo de condiciones, las personas sintetizan activamente el carbono en sus huesos de sus aminoácidos no esenciales de todas las fuentes de carbono de la dieta disponibles (hidratos de carbono, proteínas y lípidos) en lugar de a partir de una sola fuente de proteína. Dicho de otro modo, debido a que el colágeno se compone del 22% de aminoácidos esenciales y 78% de aminoácidos no esenciales, el carbono de energía de la dieta deriva en carbono hacia el colágeno, pues como señaló Schwarcz (2000, p. 208) es menos probable que el cuerpo vaya a producir los aminoácidos no esenciales si ya están presentes en la dieta. Con este principio, es posible identificar poblaciones con ingesta baja de proteínas,
así como diferencias en las fuentes de origen a partir de la relación lineal
δ13Ccol-δ13Cap. Una diferencia de 4.4‰ entre el δ13Ccol y δ13Cap, define la relación relativa del espacio entre dieta-carbonato y dieta-colágeno (De Niro y Epstein, 1978), es decir, cuando el valor de la fuente de energía es elevado en δ13C‰ en comparación con la fuente de proteína, el espacio entre el carbonato y el colágeno será mayor. Por el contrario, si el valor de la fuente de energía es bajo, el espacio entre δ13C‰ del colágeno y la apatita resulta menor. Es decir, si las fuentes de proteína animal son abundantes, las plantas que proporcionan proteínas y energía se verán reflejadas en los valores del carbonato, pero no así en los valores del colágeno, no obstante, hay que tener en cuenta que las grasas y aceites de origen marino también son una fuente principal de energía, por lo que pueden ser evidentes en los valores de la apatita (Kellner y Schoninger, 2007).
Aunque el modelo simple de regresión lineal presenta estas ventajas sobre los datos absolutos de desplazamiento, datos isotópicos en humanos arqueológicos frente a líneas de regresión obtenidas en animales de laboratorio y en su hábitat natural, revela dos limitaciones principales que se relacionan a las fuentes de proteína en grupos humanos con dietas mixtas (Froehle et al., 2010, 2012). En primer lugar, el modelo distingue poco entre fuentes de proteína C4 y marina, confundiendo la determinación de las fuentes de proteínas en poblaciones que viven en zonas costeras que albergan vegetación silvestre C4 o cultivos agrícolas. En segundo lugar, muchos individuos caen entre las líneas de proteína específica, cuyo significado en cuanto a las fuentes directas de proteínas resulta poco claro. Con el fin de resolver estas ambigüedades, se utiliza el análisis exclusivo de las variables de carbono (δ13Ccol-δ13Cap) adicionando los datos referentes a isótopos estables de nitrógeno (δ15N). Como se sabe, los valores de δ15N en colágeno óseo reflejan estrechamente las fuentes directas de proteína, ya que varían considerablemente según el consumo proporcional de diferentes tipos de plantas contra las fuentes de proteína animal, así como las fuentes de proteínas marinas contra las terrestres (De Niro y Epstein, 1981; Minagawa y Wada, 1984; Shoeninger y De Niro, 1983, 1984). Este otro modelo se basa fundamentalmente en el uso de funciones discriminantes y de conglomerados o clústers, mismos que representan de manera simultánea las tres variables para su estudio. Dos funciones representan significativamente el 98,8% de la varianza de la muestra (Froehle et al., 2012), proporcionando así un modelo confiable en donde las variables del carbono (δ13Ccol y δ13Cap) se encuentran en la primera función (F1), mientras el nitrógeno (δ15N) se encuentra representado en la segunda (F2). Para tal efecto se usa la ecuación siguiente:
Carbono: F1 = (0.322 · δ 13Capatita) + (0.727 · δ 13Ccolágeno) + (0.219 · δ 15N) + 9.354
Nitrógeno: F2 = (-0.393 · δ 13Capatita) + (0.133 · δ 13Ccolágeno) + (0.622 · δ 15N) – 8.703
De esta forma, el modelo multivariado es un método que incorpora los datos de δ13Ccol; δ13Cap y δ15N de manera holista, generando una técnica asequible para la reconstrucción de la dieta en poblaciones humanas caracterizadas por dietas mixtas. Como se puede observar, la inclusión del isótopos de nitrógeno (δ15N) resulta elemental para resolver las limitaciones de las proporciones de proteínas relacionadas del modelo de carbono de dos variables, reuniendo la información isotópica en cinco clúster o grupos alimenticios principales: 1) 100% C3 proteína/dieta total; 2) 30:70 C3:C4 dieta total, ˃50% C4 proteína; 3) 50:50 C3:C4 dieta total, proteína marina; 4) 70:30 C3:C4 dieta total, ≥65% C3 proteína; 5) 30:70 C3:C4 dieta total,˃65% C3 proteína (Froehle et al., 2012). En resumen, el empleo de ambos modelos, proporciona una base fundamentada con dietas controladas para resolver y contrastar el problema de las proporciones relativas entre fuentes de alimentos con firmas isotópicas similares, pero sobre todo, resulta de gran relevancia, para inferir patrones en las dietas en aquellas poblaciones que se encuentran conformadas principalmente por recursos de entornos costeros y con algún tipo de agricultura, diferenciando la proteína o energía total y la dieta proteica a partir de las distintas rutas metabólicas que sigue el carbono en los tejidos del esqueleto.
En este trabajo se analizan 20 muestras de colágeno y carbonatos de hueso y dientes humanos procedentes de la cueva de La Sepultura obtenidos mediante técnicas de purificación y separación (Garvie-Lock et al., 2004; Hüls et al., 2007, 2009; Sullivan y Krueger, 1981). El procedimiento previo al uso de espectrometría de masas, demandó de etapas ordenadas involucrando diferentes momentos: 1) Registro digital y en bitácora de los datos de procedencia de la muestra. 2) Corte y extracción de material con una fresa y un molino eléctrico y 3) Registro del gramaje del material antes y después de haber sido reducido a polvo de 149µm. Las muestras procesadas de los huesos y dientes fueron cuatro por cada ejemplar: 1) vial de 2 ml con 0.25 ml, para FT-IR (sin limpiar); 2) vial de 2 ml con 0.25 ml, para FTIR (limpio) para colágeno; 3) vial de 2 ml con 0.5 ml, para carbonatos y 4) tubo de 50 ml con 2.0 g, para colágeno. Una vez preparadas las muestras mediante síntesis química, se utilizó un espectrómetro de masas Finnigan Mat 253 (para muestras de esmaltes y líquidos) y al Thermo Finningan Delta plus XL (para colágeno). Los valores obtenidos, se presentan bajo los estándares y normas internacionales respecto las abundancias de los diferentes átomos que componen el carbono, el nitrógeno y el oxígeno (Coplen et al., 2002; Craig, 1953; Morales Puente y Alvarado Cienfuegos, s.f.; Morales Puente et al., 2012), mismos fueron contrastados con datos isotópicos procedentes de poblaciones de cazadores-recolectores-pescadores de la planicie costera del golfo de Texas de diversos periodos (Hard y Katzenberg, 2011). De igual modo, se conformó un perfil isotópico de fauna y flora tanto arqueológica como contemporánea de la zona, a esta última se le aplicó un factor de corrección de -1.5‰ debido al cambio climático (Wahlen, 1994). Finalmente se obtuvieron valores isotópicos de muestras de cuerpos acuíferos locales, esto con la finalidad de calcular el agua meteórica de oxígeno (δ18O) y deuterio (δ2H) específica del área de estudio, con la intención de constituir un modelo hipotético ecológico de la región. Debemos advertir que si bien, el proceso que tiene la velocidad de recambio en la que ocurren los diferentes fraccionamientos en colágeno de hueso o dentina y de bio-apatita de hueso y esmalte puede ser distinto, en este estudio se presentan en conjunto para observar su comportamiento general entre tejidos.
Resultados
Obtuvimos los valores δ de carbono 13C, nitrógeno 15N y oxígeno 18O de colágeno y apatita de 12 muestras procedentes de hueso compacto (mandíbula), 4 piezas dentarias y 4 falanges (Tabla 2). Estos resultados muestran una proporción C: N entre 2.8 a 3.8 por lo que se deduce no están afectados por diagénesis (Ambrose y De Niro, 1986; De Niro, 1985; Emery et al., 2000). El modelo hipotético del medio ambiente muestra diferencias esperadas entre las plantas C3 y las C4 junto con las CAM, lo que refleja una zona de transición entre el altiplano del norte, la sierra Madre Oriental y la Planicie costera del golfo de México, espacio donde conviven diversas especies animales y vegetales de matorral desértico espinoso, de pino-encino y selva baja caducifolia. Destaca que el maíz arqueológico se ubica cabalmente dentro de las plantas C4 con un valor δ13C de -9 (Tablas 3 y 4), valores cercanos a los reportados en diferentes poblaciones mesoamericanas y del periodo colonial en México (Montero y Nuñez Enriquez, 2011; Morales Puente et al., 2012; Warinner, 2010).
Tabla 2. Valores isotópicos de δ¹³Ccol; δ¹³Cap; δ¹5N y δ¹8O en individuos procedentes de la cueva de La Sepultura
|
Clave |
sexo |
edad |
apatita |
colágeno |
colágeno |
apatita |
apatita |
2.9 - 3.6 |
FT-IR |
|
|
|
|
|
δ¹³CVPDB |
δ¹³CVPDB (‰) |
δ15NAIR (‰) |
|
δ18OVSMOW (‰) Eq. Agua |
C/N |
%N |
%C |
|
1) TCS-mm1 |
M |
21-35 |
-2.22 |
-9.44 |
11.10 |
-3.68 |
-6.52 |
2.9 |
15.1 |
44.1 |
|
2) TCS-mm2 |
M |
21-35 |
-2.93 |
-10.97 |
11.91 |
-3.13 |
-5.96 |
2.9 |
13.9 |
40.5 |
|
3) TCS-mm3 |
M |
21-35 |
-2.85 |
-8.86 |
12.05 |
-6.90 |
-9.85 |
3.0 |
13.5 |
40.2 |
|
4) TCS-mm5 |
M |
21-35 |
-1.89 |
-9.38 |
11.82 |
-4.91 |
-7.80 |
2.9 |
15.9 |
46.5 |
|
5) TCS-mm6 |
F |
12-19 |
0.20 |
-10.10 |
12.36 |
2.38 |
-0.27 |
2.9 |
14.2 |
41.6 |
|
6) TCS-mm7 |
F |
14-16 |
-3.10 |
-10.06 |
11.68 |
-3.62 |
-6.46 |
2.9 |
14.4 |
42.0 |
|
7) TCS-mm8 |
F |
21-35 |
-3.60 |
-9.89 |
12.14 |
-4.26 |
-7.12 |
3.0 |
15.1 |
44.7 |
|
8) TCS-mm9 |
M |
21-35 |
-3.89 |
-8.65 |
12.58 |
-3.62 |
-6.46 |
3.0 |
15.1 |
44.8 |
|
9) TCS-mm10 |
F |
12-16 |
-2.70 |
-8.43 |
12.13 |
-4.80 |
-7.69 |
3.0 |
13.5 |
40.2 |
|
10) TCS-mm11 |
M |
21-35 |
-2.56 |
-10.12 |
11.84 |
-5.46 |
-8.37 |
3.0 |
15.4 |
45.6 |
|
11) TCS-mm12 |
M |
18-21 |
-4.31 |
-8.27 |
12.88 |
-3.59 |
-6.44 |
2.9 |
15.4 |
44.8 |
|
12) TCS-mm14 |
M |
21-35 |
-3.43 |
-7.87 |
13.81 |
-4.56 |
-7.44 |
2.9 |
14.6 |
42.2 |
|
13) TCS-md9 |
n/i |
Adulto |
-1.55 |
-9.42 |
12.58 |
-2.80 |
-5.62 |
2.8 |
15.1 |
42.2 |
|
14) TCS-md10 |
n/i |
Adulto |
-1.11 |
-9.21 |
14.14 |
-4.01 |
-6.87 |
2.8 |
15.2 |
42.3 |
|
15) TCS-md11 |
n/i |
Adulto |
-0.55 |
-9.24 |
13.16 |
-1.95 |
-4.74 |
2.9 |
14.5 |
41.8 |
|
16) TCS-md12 |
n/i |
Adulto |
0.31 |
-7.90 |
12.18 |
-4.09 |
-6.95 |
2.8 |
14.8 |
40.8 |
|
17) TCS-mh2 |
n/i |
Adulto |
-2.31 |
-8.59 |
12.16 |
-2.53 |
-5.33 |
2.9 |
15.2 |
44.4 |
|
18) TCS-mh3 |
n/i |
Adulto |
-3.30 |
-9.54 |
11.91 |
-1.57 |
-4.35 |
2.9 |
14.7 |
43.5 |
|
19) TCS-mh4 |
n/i |
Adulto |
-3.90 |
-9.71 |
11.58 |
-2.82 |
-5.64 |
2.9 |
14.3 |
41.3 |
|
20) TCS-mh5 |
n/i |
Adulto |
-2.12 |
-8.30 |
12.12 |
-2.21 |
-5.01 |
3.0 |
14.8 |
43.8 |
* muestras arqueológicas
Tabla 3. Valores isotópicos de δ¹³Ccol; δ¹³Cap; δ¹5N y δ¹8O de fauna procedente del área de la cueva de La Sepultura
|
Clave |
apatita |
colágeno |
colágeno |
apatita |
apatita |
2.9 - 3.6 |
|
|
δ¹³CVPDB (‰) |
δ¹³CVPDB (‰) |
δ15NAIR (‰) |
δ18OVPDB (‰) |
δ18OVSMOW (‰) Eq. Agua |
C/N |
|
1) TCT-mf1* |
-14.65 |
-19.97 |
3.65 |
-5.99 |
-8.92 |
2.95 |
|
2) TCT-mf2* |
-13.83 |
-18.31 |
4.03 |
-5.19 |
-8.08 |
2.76 |
|
3) TCT-mf3* |
-12.70 |
-18.20 |
5.47 |
-6.09 |
-9.02 |
2.75 |
|
4) TCS-mfa1 |
-15.94 |
10.62 |
- |
- |
- |
- |
|
5) TCS-mfa2 |
-19.42 |
6.04 |
- |
- |
- |
- |
* muestras arqueológicas
Tabla 4. Valores isotópicos de δ¹³C y δ¹5N de flora procedente de la cueva de la Sepultura
|
Clave |
Nombre |
Familia |
δ15NAIR (‰) |
δ13CVPDB (‰) |
δ13CVPDB (‰) |
Fotosíntesis |
|
TAP-1 |
Lechuguilla |
Asparagaceae |
6.1 |
-14.7 |
-13.2 |
C4,CAM |
|
TAP-2 |
Tuna dulce |
Cactaceae |
3.9 |
-12.7 |
-11.2 |
CAM |
|
TAP-3 |
Tuna de cardón |
Cactaceae |
4.9 |
-11.0 |
-9.5 |
CAM |
|
TAP-4 |
Cabuches |
Cactaceae |
10.1 |
-12.7 |
-11.2 |
CAM |
|
TAP-5 |
Dátil |
Asparagaceae |
5.9 |
-12.0 |
-10.5 |
C4,CAM |
|
TAP-6 |
Semilla de biznaga |
Cactacea |
4.1 |
-13.2 |
-11.7 |
CAM |
|
TAP-7 |
Fruto de tullidora |
Rhamnaceae |
6.2 |
-25.0 |
-23.5 |
C3 |
|
TAP-8 |
Pasto |
Poaceae |
0.4 |
-15.5 |
-14.0 |
C4 |
|
TAP-9 |
Hierba del cáncer |
Euphorbiacae |
4.7 |
-27.0 |
-25.5 |
C3 |
|
TAP-10 |
Hormiguilla |
Nyctaginaceae |
6.2 |
-14.4 |
-12.9 |
C4,CAM |
|
TAP-11 |
Guapilla |
Bromeliaceae |
3.6 |
-13.8 |
-12.3 |
C4,CAM |
|
TAP-12 |
Candelilla |
Euphorbiacae |
-0.6 |
-15.5 |
-14.0 |
C4,CAM |
|
TAP-13 |
Espadín |
Asparagaceae |
1.9 |
-15.9 |
-14.4 |
C4,CAM |
|
TAP-15 |
San Nicolás |
Asteraceae |
1.6 |
-28.6 |
-27.1 |
C3 |
|
TAP-16 |
Rodamundos |
Amaranthaceae |
9.5 |
-13.3 |
-11.8 |
C4,CAM |
|
TAP-17 |
Padillo |
Asparagaceae |
1.9 |
-24.7 |
-23.2 |
C3 |
|
TAP-18 |
Zacate |
Poaceae |
0.0 |
-14.7 |
-13.2 |
C4 |
|
TAP-19 |
Espadín |
Asparagaceae |
0.6 |
-12.9 |
-11.4 |
C4 |
|
TAP-20 |
para la gripa…….. |
Malvaceae |
3.2 |
-26.1 |
-24.6 |
C3 |
|
TAP-21 |
Escoba morada |
Fabaceae |
3.1 |
-27.6 |
-26.1 |
C3 |
|
TAP-22 |
Ocotillo |
Asteraceae |
4.4 |
-27.3 |
-25.8 |
C3 |
|
TAP-23 |
Salvia |
Euphorbiaceae |
4.4 |
-27.5 |
-26.0 |
C3 |
|
TAP-24 |
Guajillo |
Fabaceae |
5.7 |
-28.4 |
-26.9 |
C3 |
|
TAP-25 |
Escobilla |
Scrophulariaceae |
6.7 |
-28.7 |
-27.2 |
C3 |
|
TAP-26 |
Alfilercillo |
Geraniaceae |
6.9 |
-22.9 |
-21.4 |
C3 |
|
TAP-27 |
Ventosa |
Hydrophyllaceae |
5.2 |
-26.1 |
-24.6 |
C3 |
|
TAP-28 |
Parraleña |
Asteraceae |
2.7 |
-26.7 |
-25.2 |
C3 |
|
TAP-29 |
Espadín |
Agavaceae |
0.5 |
-14.7 |
-13.2 |
C4,CAM |
|
TAP-30 |
Hierba del zorrillo |
Serophulariaceae |
0.7 |
-27.8 |
-26.3 |
C3 |
|
TAP-31 |
Cenizo |
Serophulariaceae |
5.6 |
-26.5 |
-25.0 |
C3 |
|
TAP-32 |
Altamiz |
Asteraceae |
6.8 |
-26.9 |
-25.4 |
C3 |
|
TAP-33 |
Tabadillo |
Amaranthaceae |
9.3 |
-25.4 |
-23.9 |
C3 |
|
TAP-34 |
Cardón |
Cactaceae |
7.7 |
-12.9 |
-11.4 |
CAM |
|
TAP-35 |
Junco |
Koeberliniaceae |
5.0 |
-25.4 |
-23.9 |
C3 |
|
TAP-36 |
Sangre de grado |
Euphorbiaceae |
6.7 |
-24.1 |
-22.6 |
C3 |
|
TAP-37 |
Mezquite |
Fabaceae |
5.7 |
-24.1 |
-22.6 |
C3 |
|
TAP-38 |
Tullidora |
Rhamnaceae |
4.3 |
-27.2 |
-25.7 |
C3 |
|
TAP-39 |
Gobernadora |
Zygophyllaceae |
8.6 |
-27.1 |
-25.6 |
C3 |
|
TAP-40 |
Vizcolata |
n/i |
6.0 |
-26.9 |
-25.4 |
C3 |
|
TAP-41 |
Quiote lechuguilla |
Asparagaceae |
6.4 |
-12.9 |
-11.4 |
C4,CAM |
|
TAP-42 |
Nopal rastrero |
Cactaceae |
6.3 |
-12.0 |
-10.5 |
CAM |
|
TAP-43A |
Nopal cuija |
Cactaceae |
-0.2 |
-11.0 |
-9.5 |
CAM |
|
TAP-43B |
Nopal cuija |
Cactaceae |
6.1 |
-10.9 |
-9.4 |
CAM |
|
TAP-2C |
Pasto 1 |
Graminae |
-3.1 |
-23.0 |
-21.5 |
C3 |
|
TAP-3C |
Pasto 2 |
Graminae |
10.8 |
-10.2 |
-8.7 |
C4,CAM |
|
TAP-4C |
B3 Sotol* |
Asparagaceae |
6.9 |
-23.3 |
-21.8 |
C3 |
|
TAP-8C |
B5 Agavácea* |
Asparagaceae |
4.0 |
-24.0 |
-22.5 |
C3 |
|
TAP-10C |
B13 Semillas n/i* |
n/i |
9.4 |
-22.8 |
-21.3 |
C3 |
|
TAP-11C |
B14 Semillas n/i* |
n/i |
7.9 |
-12.0 |
-10.5 |
C4,CAM |
|
TAP-12C |
B15 Semillas n/i* |
n/i |
7.5 |
-23.6 |
-22.1 |
C3 |
|
TAP-13C |
B23 Agavácea* |
Asparagaceae |
16.0 |
-13.3 |
-11.8 |
C4,CAM |
|
TAP-15C |
B24 Hojas y varas* |
n/i |
10.3 |
-25.2 |
-23.7 |
C3 |
|
TAP-16C |
B82 Agavácea* |
Asparagaceae |
9.9 |
-13.2 |
-11.7 |
C4,CAM |
|
TAP-44 |
B84 Vaina* |
n/i |
-0.8 |
-14.1 |
-12.6 |
C4 |
|
TAP-45 |
B87 Agavácea* |
Agavaceae |
4.7 |
-15.5 |
-14.0 |
C4 |
|
TCS-mb9 |
B13/Maíz* |
Poaceae |
6.1 |
-8.97 |
- |
C4 |
* muestras arqueológicas
Como se puede observar en la Figura 4, los valores de La Sepultura se encuentran agrupados principalmente con las plantas CAM/C4 en su correspondiente nivel trófico, sin embargo, el δ15N es relativamente elevado, lo que sugiere el consumo de alimentos enriquecidos con este elemento. Lo anterior se puede deber tanto a un predominio de plantas y animales con dietas C3, C4 y CAM con altos niveles de nitrógeno, o bien, al consumo de plantas y animales que caen dentro del grupo C4 y/o marinos, insinuando con ello el manejo mixto de este tipo de recursos. Como se explicó anteriormente, la existencia de firmas isotópicas similares plantea un serio problema al momento de medir el impacto específico del medio ambiente en términos de desplazamiento isotópico. Para esclarecer lo anterior, se usaron datos de cazadores-recolectores-pescadores de la costa del golfo de Texas distribuidos en tres entornos distintos: costero, lacustre y tierra adentro con su respectivo modelo ecológico, el cual refiere principalmente a fauna (Hard y Katzenberg, 2011). Con las reservas que esto implica, sobrepusimos a estos datos un cuarto grupo que corresponde al entorno de la sierra, es decir, a la población de la cueva de La Sepultura en Tamaulipas.
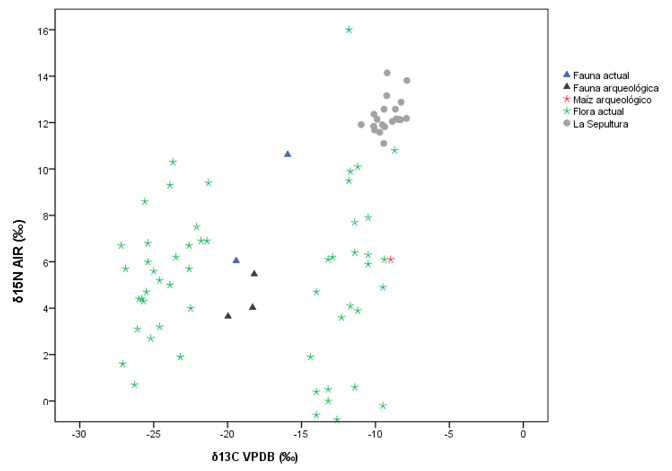
Figura 4. Valores de desplazamiento isotópico del δ13C y δ15N de la reconstrucción hipotética del medio ambiente en la sierra
Llama la atención que este grupo en cuanto a δ13C‰ presenta los valores de desplazamiento isotópico más positivos, destacando nuevamente que esta población, presenta uno de los δ15N‰ más elevados (Figura 5). Esto insinúa que están contribuyendo a su dieta alimentos enriquecidos en este elemento, es decir, vegetales y animales terrestres y probablemente recursos marinos o de agua dulce, que, por definición, presentan niveles tróficos mayores (De Niro y Epstein, 1978, 1981; Shoeninger y De Niro, 1983, 1984; Shoeninger et al., 1983). Lo anterior resulta factible si consideramos hipotéticamente, existe la viabilidad de moverse en un entorno que disponga de este tipo de recursos (Figura 6 y Figura 7). No obstante, debemos recordar que los valores isotópicos dados por diferentes tipos recursos pueden variar de región en región, por lo tanto, lo anterior también puede deberse como ya se mencionó al uso de plantas C3/CAM/C4, mismas que se encuentran enriquecidas por este elemento sobre todo en zonas semiáridas, dato apoyado por nuestra reconstrucción hipotética de flora y fauna para la zona de la sierra.

Figura 5. Valores absolutos de desplazamiento isotópico
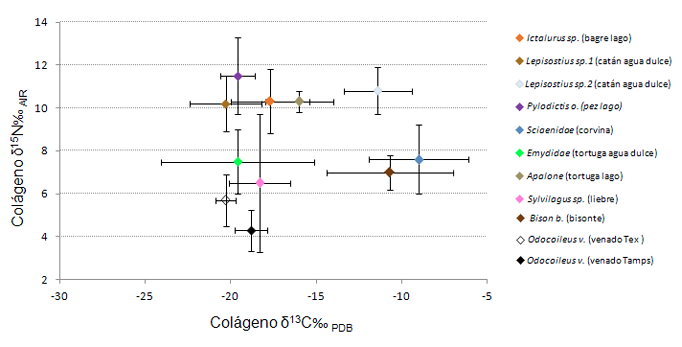
Figura 6. Valores promedio de desplazamiento isotópico en fauna de la planicie costera del golfo de Texas. Tomado y replicado de (Hard y Katzenberg, 2011)
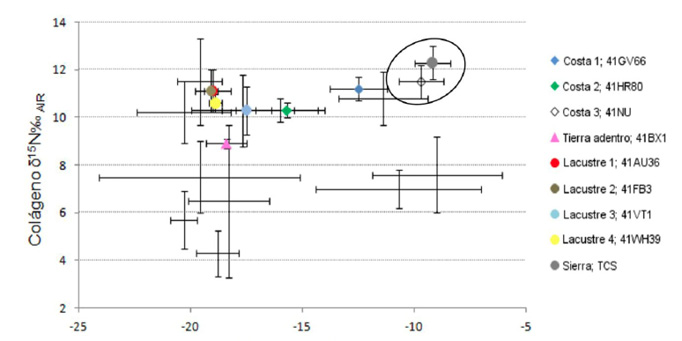
Figura 7. Valores promedio de desplazamiento isotópico de cazadores-recolectores-pescadores superpuestos a fauna de la planicie costera del golfo de Texas. Tomado y replicado de (Hard y Katzenberg, 2011)
Para exponer esta hipótesis, empleamos primero el modelo simple de regresión lineal, mismo que permite identificar diferencias entre las fuentes directas de proteína a partir de la relación δ13Ccol-δ13Cap. En primer lugar, la dispersión de La Sepultura no se comporta de manera lineal, lo que indica que las fuentes de proteína que están contribuyendo al colágeno tienen poca o nula relación con las fuentes de proteína en la apatita. De hecho, en la primera son más profusas, situando algunos sujetos mayoritariamente sobre el extremo superior de la línea específica de proteína CAM/C4, mientras otros individuos, se ajustan hacia la línea específica de proteína marina, lo que indicaría también el consumo variado de este tipo de recursos (Figura 8). Esto sugiere una tendencia hacia el uso mixto de alimentos de origen dulceacuícola o marina, que en este caso pueden proceder de cuerpos acuíferos locales como lagunas, ríos o inclusive el mar, en combinación con plantas CAM/C4 y proteína animal, lo que resultaría esperado en un entorno ecológico de transición o movilidad entre la sierra y la costa. Lo anterior plantea una rica y variedad dietética en el manejo de recursos, pues los coeficientes de correlación de Pearson corroboran que el grupo de la sierra (rp=.015, p=.949) responde a una dispersión similar a la de los grupos cazadores-recolectores-pescadores tierra adentro (rp=.034, p=.949) y lacustres o ribereños (rp=.116, p=.251), a diferencia de los de la costa (rp=.914, p=.000) los cuales si presentan regresión lineal. Para estos últimos las fuentes de proteína que contribuyen al colágeno tienen un nivel de relación mayor con las que contribuyen a la apatita (Figura 6), indicando diferencias importantes en la disponibilidad, apropiación y variedad de recursos. La explicación que se propone aquí, es que en la sierra al ser más positivos y elevados los niveles de δ13C y δ15N que el de las otras poblaciones de cazadores-recolectores-pescadores, puede deberse a un alto consumo de proteína animal terrestre, junto con el de plantas CAM/C4, probablemente a un incremento en el uso del maíz, en combinación directa con proteína acuícola o marina. De ahí su proximidad con poblaciones costeras que muestran una regresión lineal positiva, pero con una nube de dispersión característica de los grupos tierra adentro y lacustres o ribereños, las cuales su nivel de asociación es 0.
En este caso como ya vimos, las fuentes de proteína C4 y marina tienden a confundirse en la relación δ13Ccol-δ13Cap, por lo tanto, recurrimos al uso del modelo multivariado para distinguir en qué proporción la proteína animal está contribuyendo tanto al colágeno como a la bio-apatita. En La Sepultura el carbono (F1) tiene un intervalo de 3.0 a 6.4‰ con una media de 4.6‰ (s=.8, RQ=1.2, n=20), mientras para el nitrógeno (F2), un rango de -2.4 a 1.4‰ con una media de -.9‰ (s=1.1, RQ=-1.2, n=20). La posición de La Sepultura frente a los promedios de las cuatro zonas en cuestión, respecto a los cinco grupos de dietas controladas (Froehle et al., 2012), confirma que esta población
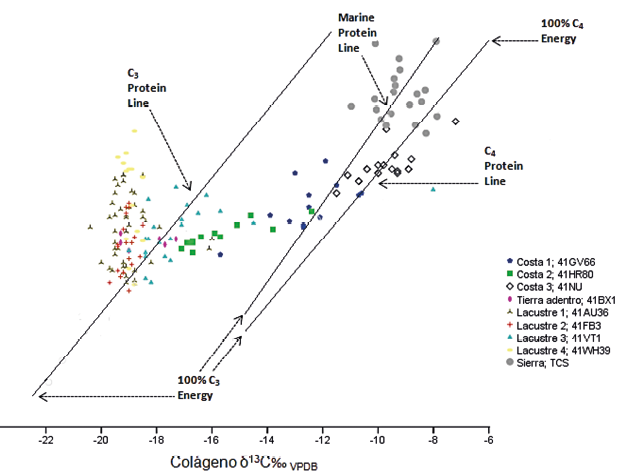
Figura 8. Modelo de regresión lineal entre δ13C del colágeno y la apatita (Kellner y Schoeninger, 2007) en las diferentes zonas de la llanura costera del golfo respecto a las de la cueva de La Sepultura
basaba su dieta en un consumo reiterado de plantas CAM/C4, plantas C3 y una proporción significativa de proteína animal terrestre e hídrica, ratificando con ello que las poblaciones más variables en sus fuentes de proteína, siguen siendo las correspondientes a las áreas tierra adentro (t=-15.419, gl=5, p˂.000), de la sierra (t=16.279, gl=19, p˂.000) y lacustres o ribereñas (t=-16.980, gl=99, p˂.000), de manera contraria a los de la costa, en la que existe asociación directa entre sus fuentes de proteína (t=6.91, gl=39, p˂.099). Como se puede apreciar en la Figura 9, el promedio de La Sepultura se sitúa al extremo derecho del Conglomerado 2 con un centroide delimitado por dos desviaciones estándar de la media; 30:70 C3: C4 dieta, ˃50% C4 proteína. Este dato ayuda a inferir, en qué proporción la alimentación de los individuos de la Cueva de La Sepultura está incorporando a su dieta aproximadamente un 70% de proteína C4 y un 30% de C3, con alrededor de un 50% o más de proteína animal. Sin embargo, en cuanto a datos individuales las muestras 5, 15 y 16 se acercan relativamente al Conglomerado 3 exhibiendo un centroide delimitado por dos desviaciones estándar de la media 50:50 C3: C4 dieta, proteína marina (Figura 10).
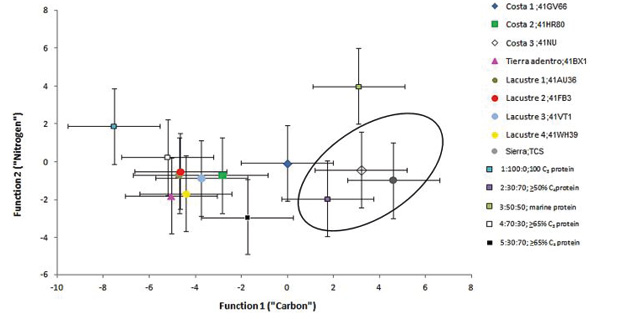
Figura 9. Modelo multivariado de funciones discriminantes (Froehle et al., 2012). La dieta de los individuos de la cueva de La Sepultura se aproxima al clúster 2; 30:70;≥50%C4proteina
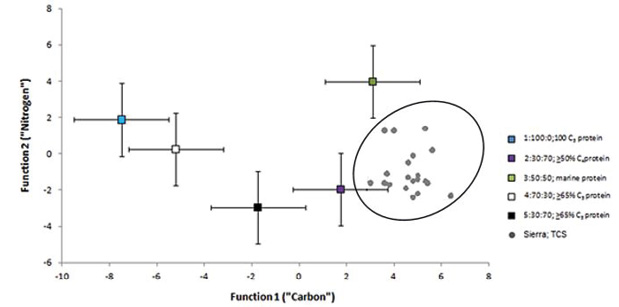
Figura 10. Modelo multivariado de funciones discriminantes (Froehle et al., 2012)
Estos individuos al parecer son los que muestran los valores más altos en δ15N, variando su dieta de manera importante de los demás, por integrar probablemente a su alimentación mayoritariamente recursos tanto de origen lacustre, ribereños o marinos, o bien, para el caso de la muestra 5, es posible que esto se deba a la influencia del nivel trófico propio de la edad del individuo. Sea como fuere, es posible plantear que los sujetos más alejados del centroide del Conglomerado 2 delimitado por dos desviaciones estándar de la media; 30:70 C3: C4 dieta, ˃50% C4 proteína, están incorporando un menor porcentaje en su dieta total fuentes de proteína C3, lo que plantea un incremento en el uso de plantas entre las que podría estar incluido el maíz y otros cultivos agrícolas o vegetación silvestre C4. Todo lo anterior resulta factible, si observamos los datos provenientes de los isótopos de δ18O, los cuales parecen indicar cierta movilidad en zonas donde existen tanto recursos de agua dulce como marinos, esto siempre y cuando, consideremos la especie de escalón y corredores que implica geográficamente el trayecto del altiplano del norte hacia la costa del golfo de México que pasa por la sierra Madre Oriental, espacios donde como se sabe para el Formativo mesoamericano, existe un desarrollo e incremento de las sociedades sedentarias y agrícolas junto con las seminómadas de cazadores-recolectores.
El promedio del agua meteórica de la región es de -6 (Figura 11), valor cercano a la mayoría de los proporcionados por los individuos de La
Sepultura, lo que indica en este caso un patrón de movilidad restringida,
pues la mayor parte se encuentran dentro del rango de variación intraindividual (Figura 12). La relación de δ18O‰ de la apatita, ayuda a inferir distintos aspectos que tienen que ver con una cierta movilidad en un entorno de transición ecológica, pues en la Figura 13 se observa que, aunque la mayor parte de los sujetos crecieron y se movieron en el entorno de la sierra, al sobreponer los datos de fauna arqueológica y contemporánea, planteamos para estos individuos, algún tipo de movilidad de una zona de mayor altitud y otro hacia un desplazamiento a nivel del mar (Figura 13).
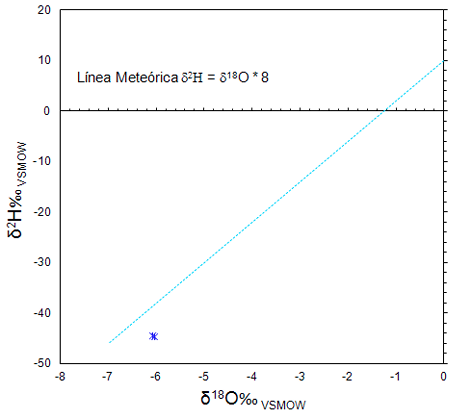
Figura 11. Línea de agua meteórica mundial de isótopos estables de oxígeno δ18O y deuterioδ2H
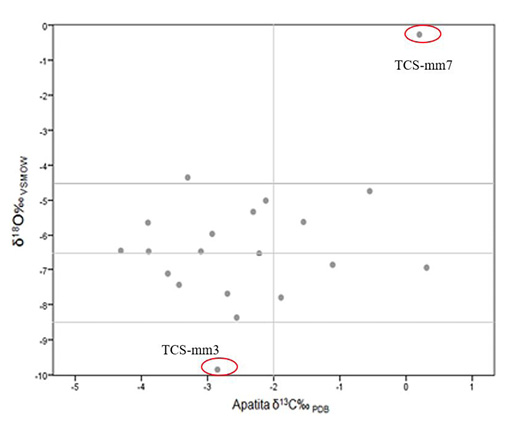
Figura 12. Valores de isótopos estables de δ18O y δ13Cap de individuos en la cueva de La Sepultura. Se señalan dos individuos que se sitúan en los extremos de mayor y menor altitud sobre el nivel del mar
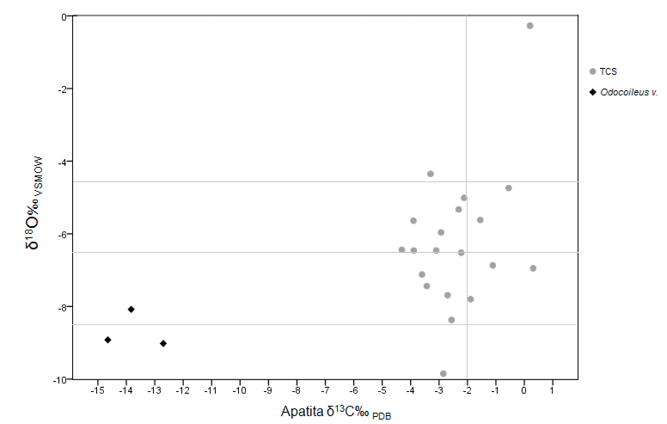
Figura 13. Valores de isótopos estables de δ18O y δ13Cap entre individuos de la cueva de La Sepultura y fauna procedente de la sierra
Discusión y conclusiones
Las técnicas isotópicas demuestran ser una herramienta eficaz para caracterizar el tipo de alimentación de las sociedades que carecen de registro etnográfico o arqueológico, así como un buen complemento de contraste en el caso de contar con este tipo de información. La extracción de elementos como el carbono, nitrógeno y oxígeno del colágeno y bio-apatita de huesos y dientes humanos, es un recurso que debe ser considerado si se pretende identificar de manera cuantitativa y diferencial, la fuente de proteína vegetal o animal terrestre o marina de las dietas del pasado. Como se ha planteado en este tipo de trabajos, las señales isotópicas de carbono impresas en colágeno y bio-apatita, contienen información acerca de un aspecto específico de la dieta, ya que se forman a través de diferentes rutas metabólicas, por lo que se asocian a diversos factores de fraccionamiento o valores de desplazamiento que pueden ser sintetizados a partir de diferentes fuentes de proteína. Las dietas bajas en proteínas, tienen menos enriquecimiento isotópico de δ15N‰ que las dietas ricas en proteínas, por lo que también en los tejidos de los consumidores con una tasa de crecimiento lento, es posible que se encuentran menos enriquecidos en este elemento que los consumidores con una tasa de crecimiento normal (Warinner, 2010). Debemos recordar que la dieta total representada por la bio-apatita, es la combinación de carbohidratos, grasas y proteínas que pueden tener cada una de las diferentes firmas isotópicas del carbono, es ingerida en cantidades diferentes por un consumidor y su composición isotópica en total de carbonos, está dada por una ecuación de regresión lineal (Schwarcz, 2000).
En este estudio, las diferencias de fraccionamiento isotópico de δ13C mediante las distintas rutas que sigue el metabolismo, así como del δ15N y δ18O del colágeno y bio-apatita de muestras procedentes de la cueva de La Sepultura, hacen suponer que los recursos obtenidos y aprovechados por esta población, reflejan una parte del complejo proceso de interface en el tema de la adopción o aparición de la agricultura en la zona. Esta problemática, debe en todo caso ser analizada no como un proceso abrupto entre dos formas distintas de aprovechamiento o producción de recursos, sino más bien, como un proceso lento, paulatino, divergente y quizás optativo, que implicó otra serie de transformaciones adaptativas en las condiciones de vida y salud, sociales, demográficas y culturales en la manera de cómo el hombre se desplaza, o mantiene en un lugar fijo para apropiarse de los recursos disponibles en el medio ambiente.
En este caso particular, las características morfológicas y culturales de los restos óseos de este trabajo, sugieren se trata de un grupo con características propias de los así denominados por la arqueología como cazadores-recolectores-pescadores del Norte de México, sin embargo, su perfil isotópico, en cierto modo, configura una tendencia al de grupos agrícolas de Mesoamérica (Arnaud Salas, 2014; Morales Puente et al., 2012; Somerville et al., 2013), sin que con ello quiera justificarse un proceso de aculturación de tipo difusionista, sino más bien, la apreciación de estrategias diversas y particulares relacionadas con su medio ambiente y tipo de organización social. En este sentido, aunque si se puede decir que durante el proceso de adaptación de los cultivos estacionales éste fue cobrando cada vez mayor relevancia, sobre todo en lo que respecta al periodo de ocupación de la cueva para la fase Mesa del Guaje (1650-1050 a.C) o fase Venadito I (1000-200 a.C), comienza también a vislumbrarse el inicio y apogeo de sociedades semi-sedentarias agrícolas que no abandonan la caza y la recolección, por lo que estamos frente una dieta rica y variada, que junto con otras manifestaciones culturales específicas que incluyen aldeas con arquitectura de piedra y tierra, aparición de la cerámica y variedad de figurillas, dan lugar a una serie de elementos contrastantes. Esto último es interesante de analizar, pues paralelamente a lo que ocurre con las poblaciones sedentarias en la cuenca baja del Pánuco, que pudieron tener contacto con estos grupos, es sabido que desde el 1600 a.C., ya existen aldeas permanentes sobre todo hacia la zona Sur del estado en la región mejor conocida como la Huasteca (Merino Carrión y García Cook, 2002, 2004), siendo de nuestro mayor interés preguntarse qué está ocurriendo con la diversidad de las poblaciones en la zona serrana durante los inicios del Formativo.
Con la información presentada en este trabajo, planteamos que las poblaciones antiguas del Suroeste de Tamaulipas, reflejan isotópicamente una parte del proceso de interface o adaptación de los cultivos estacionales de baja intensidad a la de una mayor actividad agrícola, que se ve consolidada, con el surgimiento, apogeo e interacción de importantes complejos culturales tanto en la zona de la costa con los Huastecos, en la sierra con los Pueblito y el altiplano–zona media potosina con la región de Río Verde/Pame. A nivel regional, podemos suponer que se trata de grupos que vienen aportando elementos culturales y poblacionales desde el Norte de México, lo que indica una liminalidad con la denominada frontera septentrional mesoamericana. La población de la cueva de La Sepultura entonces, nos habla de grupos seminómadas (Arias López y Velasco González, 2016) agricultores de baja intensidad con un perfil biológico y cultural característico de la tradición del Desierto, la cual tiene importante presencia en todo el altiplano del norte cuando menos desde el Arcaico medio al Prehistórico tardío (1000 a.C.-1500 d.C.). En este sentido, de alguna manera están coexistiendo o en su caso, están estrechamente vinculados con las poblaciones de agricultores semi-nómadas del Formativo temprano o finales del Arcaico medio en el noreste de México. Esta postura, mantiene el supuesto de que más que hablar de un detrimento en el proceso de apropiación de recursos interpretado a partir de la correlación lineal debida a una baja ingesta de proteína animal, por un incremento en la ingesta de proteína C4, tenemos una rica y variada alimentación que caracteriza a poblaciones con una dieta mixta. Estos hallazgos, nos hacen preguntarnos, por qué si en esta fase de desarrollo cualitativo de estas sociedades se tiene una disponibilidad amplia de recursos que son adecuadamente aprovechados por el organismo, en el registro arqueológico, algunos otros restos óseos humanos procedentes de diferentes sitios prehispánicos del Formativo al Clásico, comienzan a mostrar un detrimento en sus condiciones de vida y salud (Velasco González, 2019).
Agradecimientos
Este trabajo fue posible gracias al apoyo técnico y financiero de la Universidad Nacional Autónoma de México y del Instituto Nacional de Antropología e Historia. De igual modo nuestro más profundo agradecimiento al doctor Arturo Mora Olivo del Instituto de Ecología Aplicada de la Universidad Autónoma de Tamaulipas (UAT) y las comunidades de Joyas de Berrendo y Maravillas del ejido Lázaro Cárdenas, así como al ayuntamiento de Tula y al gobierno del Estado de Tamaulipas quienes en todo momento otorgaron las facilidades necesarias para la realización de este trabajo de investigación.
Bibliografía
Adriano-Morán, C. C. y Martínez Yrízar, D.
(s.f.). Análisis preliminar de los materiales botánicos de la cueva de La Sepultura, Tamaulipas, México. Instituto de Investigaciones Antropológicas.
Ambrose, S.
(1993). Isotopic analysis of paleodiets: Methodological and interpretative considerations. En M. K. Sandford (Ed.), Investigations of ancient human tissue: Chemical analysis in anthropology (pp. 59-121). Gordon and Breach Science.
Ambrose, S. y De Niro, M.
(1986). Reconstruction of African human diet using bone collagen, carbon and nitrogen isotopes ratios. Nature, (319), 321-324.
Arias López, J. M. y Velasco González, J. E.
(2016). Microadaptación de grupos paleoamerindios en el noreste de México. Dinámica funcional del fémur y su relación con las estrategias de subsistencia. En L. C. Jiménez, S. C. Serrano, C. B. Valle, A. F. J. Aguilar, G. A. González y C. López (Eds.), El poblamiento temprano en América (pp. 73-102). Museo del Desierto. Instituto Nacional de Antropología e Historia.
Arnaud Salas, M.
(2014) Procedencia y dieta de una muestra ósea procedente de la Ventilla 92-94 Teotihuacan. Universidad Nacional Autónoma de México.
Caro Gómez, J. A., Álvarez García, G., Cámara Artigas, R., Molina Rodríguez, J., Velasco González, J., Silva Cárdenas, T. y Pérez Silva, C. V.
(2012) Estudio geoarqueológico de cuevas mortuorias en Tamaulipas; Análisis e interpretación de los restos materiales, bioculturales y paleoambientales. Resultados preliminares de la campaña 2011-2012. Ministerio de Educación, Cultura y Deporte, España.
Coplen, T. B., Hopple, J. A., Böhlke, J. K., Peiser, H. S., Rieder, S. E., Krouse, K. J., Rosman, K. J. R., Ding, T., Vocke Jr., R. D., Révész, K. M., Lamberty, A., Taylor, P. y De Biévre, P.
(2002) Compilation of minimum and maximum isotope ratios of selected elements in naturally occurring terrestrial materials and reagents. United States Department of the Interior. United States Geological Survey.
Craig, H.
(1953) The geochemistry of the stable carbon isotopes. Geochimica and Cosmochimica Acta, (3), 53-92.
Daux, V., Lecuyer, C., Herán, M., Amiot, R., Simon, L., Fourel, F., Adam, F., Lynnerup, N. y Reychler, H.
(2008) Oxygen isotope fractionation between human phosphate and water revisted. Journal of Human Evolution, 55 (6), 1138-1147.
De Niro, M.
(1985) Post-mortem preservation and alteration of in vivo bone collagen isotope ratios in relation to paleodiatary reconstruction, Nature, 317, 806-809.
De Niro, M. y Epstein, S.
(1978) Influence of diet on the distribution of carbon isotopes in animals. Geochimica and Cosmochimica Acta, (42), 495-506.
(1981) Influence of diet on the distribution of nitrogen isotopes in animals. Geochimica and Cosmochimica Acta, (45), 341-351.
Delgado, A., Iacumin, P., Stenni, B., Sánchez, B. y Longinelli, A.
(1995) Oxygen isotope variations of phosphate in mammalian bone and tooth enamel. Geochimica and Cosmochimica Acta, 59 (20), 4299-4305.
Ekholm, G. F.
(1943) Relations between Middle America and the Southeast. En El norte de México y el sur de Estados Unidos, III Mesa Redonda de la Sociedad Mexicana de Antropología, (pp. 276-282).
(1944) Excavations at Tampico-Panuco in the Huasteca, México. Anthropologists papers of the Natura History Museum, XXXVIII (5), 321-509.
Emery, K. F., Wright, L. E. y Shwarcz, H.
(2000) Isotopic analysis of ancient deer bone: Biotic stability in collapse period Maya land use. Journal of Archaeological Science, (27), 537-550.
Eubanks, M. W.
(2001) The mysterious origin of maize. Economic Botany, 55 (4), 492-514.
Flannery, K.
(1973) The origins of agriculture. Annual Review of Anthropology, 2, 271-310.
Froehle, A. W., Kellner, C. M. y Shoeninger, M. J.
(2010) FOCUS: effect of diet and protein source on carbon stable isotope ratios in collagen: follow up to Warinner and Tuross (2009). Journal of Archaeological Science, (37), 2662-2670.
(2012) Multivariate carbon and nitrogen stable isotope model for the reconstruction of prehistoric human diet. American journal of physical anthropology, (147), 352-359.
Garvie-Lock, S., Varney, T. y Katzenberg, M.
(2004) Preparation of bone carbonate for stable isotope analysis: The effects of treatment time and acid concentration. Journal of Archaeological Science, 31, 763-776.
Gat, J. R.
(1996) Oxygen and hydrogen isotopes in the hydrologic cycle. Annual Review of Earth and Planetary Sciences, 24 (1), 225-262.
Hanselka, K.
(2008) Las cuevas de Ocampo en contexto: Investigación sobre el desarrollo del cultivo de plantas en el suroeste de Tamaulipas. Instituto Nacional de Antropología e Historia.
(2011) Prehistoric Plant Procurement, Food Production, and Land Use in Southwestern Tamaulipas, Mexico. Washington University in St. Louis.
Hard, R. J. y Katzenberg, M. A.
(2011) Stable isotope study of hunter-gatherer-fisher diet, mobility and intensification on the Texas Gulf Coastal Plain. American Antiquity, 76 (4), 709-751.
Harlan, J. R.
(1971) Agricultural Origins: Centers and Noncenters. Science, 174 (4008), 468-474.
Hole, F.
(1993) The origins of agriculture and settled life. Journal of Field Archaeology, 20 (3), 376-378.
Hughes, J. T.
(1947) An Archaeological Reconnaissance in Tamaulipas, Mexico. American Antiquity, 13 (1), 33-39.
Hüls, M., Grootes, P. y Nadeau, M.
(2007) How clean is ultrafiltration cleaning bone collagen?. Radiocarbon, 49, 193-200.
(2009) Ultrafiltration: Boon or bane?, Radiocarbon, 51, 613-625.
Jim, S., Ambrose, S. y Evershed, R. P.
(2004) Stable carbon isotopic evidence for differences in the dietary origin of bone cholesterol, collagen, and apatite: implications for their use in paleodietary reconstruction. Geochimica and Cosmochimica Acta, (68), 61-72.
Kellner, C. M. y Schoeninger, M. J.
(2007) A simple carbon isotope model for reconstructing prehistoric human diet. American Journal of Physical Anthropology, 133 (4), 1112-1127.
Kirchhoff, P.
(1943a) El Norte de México y el sur de Estados Unidos. En III Mesa Redonda sobre los problemas antropológicos de México y Centroamérica. Sociedad Mexicana de Antropología.
(1943b) La unidad básica de la cultura de los recolectores-cazadores del Norte de México, Sociedad Mexicana de Antropología.
(1954) Gatherers and Farmers in the Greater Southwest: A Problem in Classification. American Anthropologist, 56 (4), 529-550.
Krieger, Alex D.
(1945) An inquiry into supposed mexican influence on a prehsitoric cult in the southern United States. American Anthropologist, 47, 483-515.
Krueger, H. W. y Sullivan, C. H.
(1984) Models for carbon isotope fractionation between diet and bone. En J. Turlund y P. Johnson (Eds.), Stable isotope in nutritions (pp. 205-220). American Chemical Society Symposium Series.
Lee-Thorp, J. A., Sealy, J. C. y Van der Merwe, N. J.
(1989) Stable carbon isotope ratio differences between bone collagen and bone apatite, and their relationships to diet, Journal of Archaeological Science, 16, 585-599.
Lyne, B. S., Cox, G. y Sealy, J.
(2001) Determining isotope life history trajectories using bone density fractionation and stable isotope mesurements: A new approach, American journal of physical anthropology, 116, 66-79.
MacNeish, R. S.
(1947) A Preliminary Report on Coastal Tamaulipas, Mexico. American Antiquity, 13 (1), 1-15.
(1949) Prehistoric relationships between the cultures of the Southeastern United States and Mexico in the light of an archaeological survey of the State of Tamaulipas, Mexico [Tesis de doctorado, University of Chicago]. https://www.proquest.com/openview/97fb6281f9e92c7248db5086e614ff3e/1?pq-origsite=gscholar&cbl=18750&diss=y
(1950) A synopsys of archaeological sequence in the Sierra de Tamaulipas. Revista mexicana de estudios antropológicos, XI, 79-96.
(1954) An early archaeological site near Pánuco, Veracruz. Transactions of American Phylosophical Society.
(1958) Preliminary archaeological investigations in the Sierra de Tamaulipas, México. Transactions of the American Philosophical Society.
(1998) Preliminary excavation in the Ocampo, Tamaulipas, Mexico. Philadelphia.
(2001) A response to long´s radicarbon determinations that attempt to put acceptable cronology on the fritz. Latin American Antiquity, 12 (1), 99-104.
Mangelsdorf, P. C., MacNeish R. S. y Galiant, W. C.
(1964) Domestication of Corn. Science, 143 (3606), 538-545.
Merino Carrión, L. y García Cook, A.
(1987) Proyecto arqueológico Huasteca, Arqueología, (1), 31-72.
(1997) Secuencia cultural para el Formativo en la cuenca baja del río Pánuco, Arqueología, 32, 5-27.
(2002) El Formativo temprano en la cuenca baja del río Pánuco: Fases Chajil y Pujal. Arqueología, (28), 49-74.
(2004) Secuencia cultural para el Formativo en la Cuenca baja del río Pánuco. Arqueología, 32, 5-27.
Minagawa, M. y Wada, E.
(1984) Stepwise enrichment of 15N along food chains: further evidence and the relation between δ15N and animal age. Geochimica and Cosmochimica Acta, 48, 1135-1140.
Montero, C. y Núñez Enríquez, L. A.
(2011) Salud y dieta entre los entierros de Chinnikihá. Primeros resultados.Estudios de antropología biológica, XV: 139-166.
Morales Puente, P. y Alvarado Cienfuegos, E.
(s.f.) Metrología de isótopos estables y materiales de referencia utilizados para la determinación isotópica de carbono, nitrógeno, oxígeno, hidrógeno y azufre. Instituto de Geología, Universidad Nacional Autónoma de México.
Morales Puente, P., Cienfuegos Alvarado, E., Manzanilla Naim, L. y Otero Trujano, F.
(2012) Estudio de la paleodieta empleando análisis de isótopos estables de los elementos carbono, oxígeno y nitrógeno en restos humanos y fauna encontrados en el barrio teotihuacano de Teopancazco, Teotihuacan. En L. R. Manzanilla (Ed.), Estudios arqueométricos del centro de barrio de Teopancazco en Teotihuacan (pp. 347-423). Instituto de Investigaciones Antropológicas, Universidad Nacional Autónoma de México.
Nier, A. O. y Gulbransen, E. A.
(1939) Variations in the relative abundance of the carbon isotopes. Journal of the American Chemical Society, 61, 697-698.
Pérez Silva, C. V., Cárdenas Silva, T., González Velasco, J. E., Rocha Hernández J. A. y García Vázquez, G.
(2010) Informe de campo del rescate arqueológico La Cueva de la Calavera; Tula, Tamaulipas. Instituto Nacional de Antropología e Historia, Ciudad Victoria, Tamaulipas.
Pérez Silva, C., Cárdenas Silva, T., González Velasco, J. E., Meza Peñaloza, A. y Serrano Sánchez, C.
(2011a) Estudio biocultural en restos óseos humanos procedentes de cuevas mortuorias en Tamaulipas (Informe Técnico 1). Instituto Nacional de Antropología e Historia.
(2011b) Proyecto Estudio biocultural de restos óseos humanos procedentes de cuevas mortuorias en Tamaulipas. Instituto Nacional de Antropología e Historia.
Rodriguez Loubet, F.
(1985) Les chichimeques. Centro de Estudios Mexicanos y Centroamericanos.
Salinas Rivera, M.
(2012) Indígenas del delta del río Bravo; Su papel en la historia del sur de Texas y el noreste de México. Universidad Autónoma de Tamaulipas.
Schwarcz, H. P.
(2000) Some biochemical aspects of carbon isotopic paleodiet studies. En S. Ambrose y M. Katzenberg (Eds.), Biogeochemical approaches to paleodietary analysis. Kluwer Academic.
Shoeninger, M. J. y De Niro, M.
(1983) Stable nitrogen isotope ratios of bone collagen reflect marine and terrestrial components of prehistoric human diets. Science, 220, 1381-1383.
(1984) Nitrogen and carbon isotopes compositions of bone collagen from marine and terrestrial animals. Geochimica and Cosmochimica Acta, 48, 625-639.
Shoeninger, M. J., De Niro, M. y Tauber, H.
(1983) Stable nitrogen isotope ratios of bone collagen reflect marine and terrestrial components of prehistoric human diets. Science, 220, 1381-1383.
Smith, Bruce
(1997) Reconsidering the Ocampo Caves and the Era of Incipient Cultivation in Mesoamerica. Latin American Antiquity, 8 (4), 342-383.
(1998a) Between foraging and farming. Science, 279, 1651-1671.
(1998b) The emergence of agriculture. Scientific American Library.
(2001) Low-level food production. Journal of Archaeological Research, 9 (1), 1-43.
Somerville, A., Fauvelle, M. y Froehle, A.
(2013). Applying new approaches to modeling diet and status: isotopic evidence for commoner resiliency and elite variability in the Classic Maya lowlands. Journal of Archaeological Science, 40, 1539-1553.
Stresser-Pean, G.
(1977). San Antonio Nogalar, La Sierra de Tamaulipas et la frontière Nord-est de la Mésoamérique. Misión arqueológica y etnológica francesa a México.
Sullivan, C. H. y Krueger, H. W.
(1981). Carbon isotopes analysis of separate chemical phases in modern and fossil bone. Nature, 292, 333-335.
Tesch, M.
(1991). El área de Alaquines: una zona de contactos. En M. A. Cabrero (Ed.), II Coloquio Pedro Bosch Gimpera (pp. 443-459). Instituto de Investigaciones Antropológicas.
(1996). Aspectos culturales en el área central de la zona media potosina. En L. Torre (Ed.), Xi’oi Coloquio pame. Los pames de San Luis Potosí y Querétaro. Centro de Investigaciones Históricas de San Luis Potosí.
Tieszen, L. y Fagre, T.
(1993). Effect of diet quality and composition on the isotopic composition of respiratory CO2, bone collagen, bioapatite, and soft tissues. En J. Lambert y G. Grupe (Eds.), Prehistoric human bone: archaeology at the moleculr level (pp.121-156). Springer-Verlag.
Van der Merwe, Nikolaas J y John C Vogel
(1978) 13C content of human collagen as a measure of prehistoric diet in woodland North America. Nature, 276 (5690):815-816.
Velasco González, J. E.
(2011). Limpieza, restauración y catalogación de restos óseos humanos procedentes del rescate arqueológico; La Cueva de la Calavera, Tula, Tamaulipas (Informe). Instituto Nacional de Antropología e Historia.
(2016). Tafonomía e isótopos estables en la cueva de la Sepultura (1200 a. C.) Tula, Tamaulipas. Universidad Nacional Autónoma de México.
(2019). Enterramientos humanos del sitio arqueológico Lomas del Real, Altamira, Tamaulipas; Condiciones de vida y salud durante el Formativo terminal (300 a. C. - 200 d. C.) en el Norte de la Huasteca. Instituto Nacional de Antropología e Historia.
Velasco González, J. E., Silva Cárdenas, T., Pérez Silva, C. V., Meza Peñaloza, A. y Serrano Sánchez, C.
(2013). Análisis de Materiales de la Temporada I Cuevas Morturias de Tamaulipas (Informe técnico). Instituto Nacional de Antropología e Historia.
Vogel, G. C. y N. J. Van der Merwe
(1977) Isotopic evidence for early maize cultivation in New York State. American Antiquity, 42:238-242.
Wahlen, M.
(1994). Carbon dioxide, carbon monoxide and methane in the atmosphere: abundance and isotopic composition. Stable isotopes in ecology and environmental science, 93-113. [Editorial?]
Warinner, G. C.
(2010). Life and death in Teposcolula, Yucundaa: Mortuary, archeogenetic and isotopic investigations of the Early Colonial Period in México [Tesis de doctorado, Harvard University]. http://christinawarinner.com/wp-content/uploads/2017/07/Warinner_Dissertation_June252010-2.pdf
William, Merrill I, Robert J Hard, Jonathan B Mabry, Fritz J Gayle, Karen R Adams, John R Roney y A. C MacWilliams
(2009) The difussion of maize to the southwestern United States and its impact. PNAS, 105 (50), 21019-21026.